Why Do Europeans Have So Many Hair and Eye Colors?
Université Laval (Canada) and St. Andrews
University (Scotland).
[This article summarizes Frost (2006) and includes updates in response to criticism.]
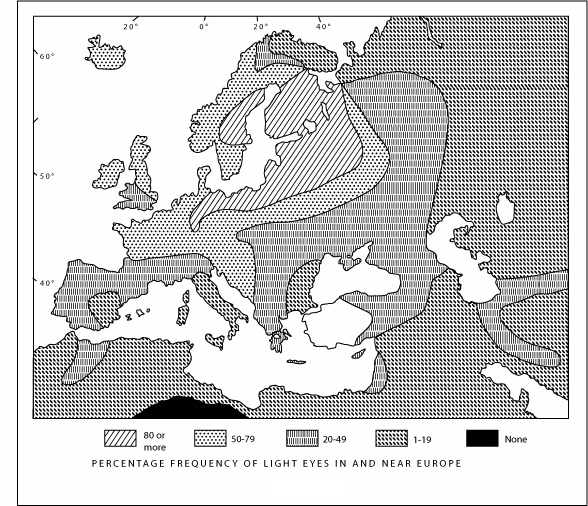
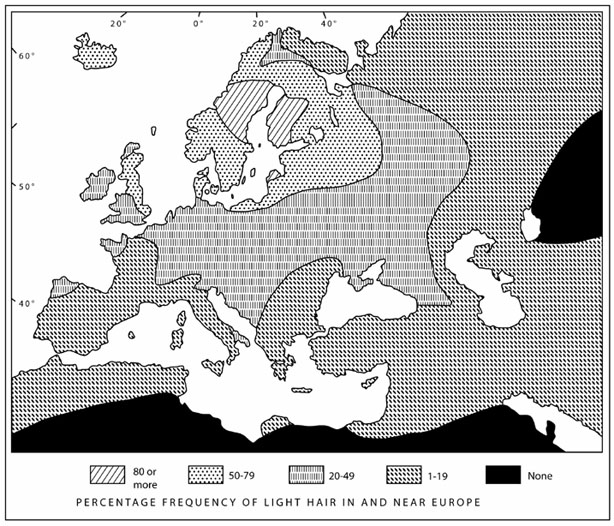 Most humans have only one hair color and one eye color.
Europeans are a big exception: their hair is black but also
brown, flaxen, golden, or red; their eyes are brown but also
blue, gray, hazel, or green. This diversity reaches a maximum in
an area centered on the East Baltic and covering northern and
eastern Europe. If we move outward, to the south and east, we see
a rapid return to the human norm: hair becomes uniformly black
and eyes uniformly brown.
Most humans have only one hair color and one eye color.
Europeans are a big exception: their hair is black but also
brown, flaxen, golden, or red; their eyes are brown but also
blue, gray, hazel, or green. This diversity reaches a maximum in
an area centered on the East Baltic and covering northern and
eastern Europe. If we move outward, to the south and east, we see
a rapid return to the human norm: hair becomes uniformly black
and eyes uniformly brown.
Why this color diversity? And why only in Europe? Some believe it to be a side effect of natural selection for fairer skin to ensure enough vitamin D at northern latitudes. Yet skin color is weakly influenced by the different alleles for hair color or eye color apart from the ones for red hair or blue eyes. Some have no effect at all on skin pigmentation.
Others put the cause down to intermixture with Neanderthals. Yet, according to the mtDNA that has been retrieved, no genetic continuity is discernible between late Neanderthals and early modern Europeans. Perhaps there was some gene flow between the two groups, but certainly not enough to account for the large number of Europeans with neither black hair nor brown eyes.
For others still, this color diversity arose through random factors: genetic drift, founder effects, relaxation of natural selection, etc. But these factors could not have produced such a wide variety of hair and eye hues in the 35,000 years that modern humans have inhabited Europe. The hair-color gene (MC1R) has at least 7 alleles that exist only in Europe and the same is probably true for the eye-color gene (OCA2). If we take the hypothesis of a relaxation of selection, nearly a million years would be needed to accumulate this amount of diversity. Moreover, it is odd that the same sort of diversification has evolved at two different genes whose only point in common is to color a facial feature.
Thus, some kind of non-random process seems
to have targeted hair and eye color per se, that is, as
visible characteristics. But how? And why? For some, including
the geneticist Luigi L. Cavalli-Sforza, the answer is sexual
selection. This mode of selection intensifies when males
outnumber females among individuals ready to mate, or vice versa.
The sex in excess supply has to compete for a mate and resorts to
the same strategies that advertisers use to grab attention, such
as the use of bright or striking colors.
 Guppy males (Poecilia reticulata) caught on a single morning from a single creek (Brooks 2002). Rare-color advantage has been studied mainly in guppies and fruit flies but also occurs in other animals. In addition, a number of bird species exhibit color polymorphisms for which the mode of selection remains unclear. Whatever the cause, color polymorphisms are relatively uncommon. They are often hindered by two evolutionary constraints: 1) high predation pressure, this being a constraint on color traits in general, and 2) presence of related species within the same geographic range, apparently because too much intraspecific variability makes it harder to recognize one's own species and leads to hybridization.
Representative eye colors (Sturm and Frudakis 2004) |

In other animals, bright colors are usually due to sexual selection. Sometimes the result may be a "color polymorphism" (see box). A potential mate will respond not simply to a bright color but also to a rare one that stands out from the crowd. By enhancing reproductive success, however, such a color will also become more common and less eye-catching. Sexual attraction will then shift to less common variants, the eventual result being an equilibrium that maximizes color diversity.
This sort of rare-color advantage has been reported in humans. An American researcher, Thomas Thelen, prepared three series of slides featuring attractive women: one with 6 brunettes; another with 1 brunette and 5 blondes; and a third with 1 brunette and 11 blondes. Male subjects then had to select the woman in each series they would most prefer to marry. For the same brunette, preference increased significantly from the first to the third series, i.e., in proportion to the rarity of the brunettes. This rare-color preference may account for the wide range of human hair and eye phenotypes we see today.
But why is hair and eye color so much more diverse in Europe than elsewhere? Perhaps because sexual selection was much stronger among ancestral Europeans than in other human populations.
Sexual selection intensifies when the "Operational Sex Ratio" (OSR) ceases to be balanced, i.e., when too many of one sex are competing for too few of the other. To understand why this may have happened in ancestral humans, we can examine the demography of present-day hunter-gatherer bands. Such groups usually develop an OSR imbalance for two reasons: 1) hunting distances are longer and have increased the death rates of young men, typically because game animals are more mobile and/or less numerous per unit of land area; and 2) the cost of providing for a second wife is higher and has reduced the incidence of male polygamy (polygyny), typically because women are procuring less food for themselves through food gathering. As a rule, OSRs are less balanced further away from the equator. In the Temperate Zone, and even more so in the Arctic, game animals roam over larger territories and gatherable food is less available in winter.
The most extreme OSR imbalance occurs among hunting peoples of the "steppe-tundra," where almost all consumable biomass is in the form of highly mobile and spatially concentrated herbivores such as caribou, reindeer, or muskox. On the one hand, men die younger because they have to cover long distances in search of herds, with no alternate food sources. On the other, men are less polygynous because they bear almost the full cost of feeding their families in a habitat that offers women little opportunity for food gathering. With fewer men altogether and even fewer polygynous ones, women have to compete for a limited supply of potential husbands.
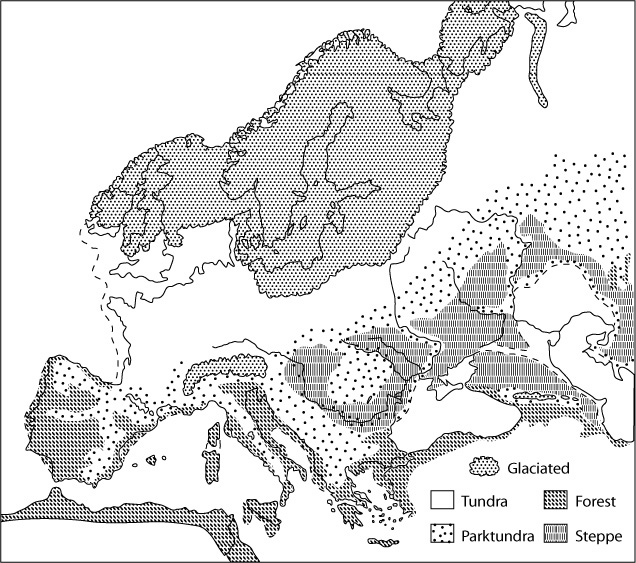 Ecological zones
in Europe at the last glacial maximum, ca 18,000 BP. Ecological zones
in Europe at the last glacial maximum, ca 18,000 BP. |
Steppe-tundra is now reduced to fragments along the northern fringes of Eurasia and North America. During the last ice age, however, when modern humans first arrived, the Scandinavian icecap had pushed it farther south onto the plains of Europe. The more intense sunlight, combined with fertile loess soils, created an expanse of steppe-tundra with unusually high bioproductivity, even at the peak of the ice age. Less productive was the Asian steppe-tundra east of the Urals. It was drier, farther north, and largely polar desert, especially at the glacial maximum. Prospects were better for continuous and substantial human settlement on the European portion of this ecological zone.
The European steppe-tundra was distinctive in another way. It took in an area that covers almost the same area where hair and eye color is today most diverse. Could this be an imprint left on the human genetic landscape by sexual selection?
Perhaps. But more proof is needed. One tantalizing piece of evidence is the possibility that these new hair and eye colors are mildly sex-linked, as would be expected if women were more strongly selected for such characteristics. According to an unpublished British study, non-black-haired and non-brown-eyed individuals have longer second fingers in relation to their fourth fingers. This indicates that the new hair and eye colors are associated with a higher ratio of estrogen to testosterone before birth. Interestingly, blond hair has arisen independently among some Aborigines of central Australia and is more frequent there in women than in men.
The Aborigine example points to another avenue for research: populations outside Europe among whom new hair and eye colors seem to have appeared independently. There are a few such cases: blond hair among central Australian Aborigines, brown hair among the Yukhagir of eastern Siberia, and fair hair among some Inuit bands of the western Canadian Arctic. Is hair color less diverse in such populations than in Europeans because sexual selection has been less intense or has acted over a shorter period of time?
A final avenue for research might be to extract DNA from skeletal remains in order to chart European MC1R and OCA2 variability over the last ice age. If the sexual selection hypothesis is true, MC1R and OCA2 variability should have developed almost entirely during this time window (c. 25,000 - 10,000 BP).
References:
Abbie, A.A., and W.R. Adey. 1953. Pigmentation in a central Australian tribe with special reference to fair-headedness. American Journal of Physical Anthropology 11:339-359.
Brooks, R. 2002. Variation in female mate choice within guppy populations: population divergence, multiple ornaments and the maintenance of polymorphism. Genetica 116:343-358.
Duffy, D.L., N.F. Box, W. Chen, J.S. Palmer, G.W. Montgomery, M.R. James, N.K. Hayward, N.G. Martin, and R.A. Sturm. 2004. Interactive effects of MC1R and OCA2 on melanoma risk phenotypes. Human Molecular Genetics 13:447-461.
Frost, P. 2006. European hair and eye color - A case of frequency-dependent sexual selection? Evolution and Human Behavior 27:85-103
Hughes, K.A., L. Du, F.H. Rodd, and D.N. Reznick. 1999. Familiarity leads to female mate preference for novel males in the guppy, Poecilia reticulata. Animal Behaviour 58:907-916.
Makova, K, and H. Norton. 2005. Worldwide polymorphism at the MC1R locus and normal pigmentation variation in humans. Peptides 26:1901-1908.
Sturm, R.A., and T.N. Frudakis. 2004. Eye colour: portals into pigmentation genes and ancestry. Trends in Genetics 20:327-332.
Thelen, T.H. 1983. Minority type human mate
preference. Social Biology, 30, 162-180.